 题目内容
(请给出正确答案)
题目内容
(请给出正确答案)
[主观题]
M.Green在果蝇中发现了一个特殊的X连锁的焦刚毛(sn)基因。雌性纯合体具有焦刚毛,但在它们的头、胸、
M.Green在果蝇中发现了一个特殊的X连锁的焦刚毛(sn)基因。雌性纯合体具有焦刚毛,但在它们的头、胸、腹部有许多区域具有sn+(野生型)刚毛。该雌性纯合体果蝇与sn雄性杂交时,一些雌性亲本仅产生焦刚毛后代,而另一些雌性亲本则以可变的频率产生焦刚毛和野生型的后代。试解释之。
查看答案

 如果结果不匹配,请 联系老师 获取答案
如果结果不匹配,请 联系老师 获取答案

 更多“M.Green在果蝇中发现了一个特殊的X连锁的焦刚毛(sn)…”相关的问题
更多“M.Green在果蝇中发现了一个特殊的X连锁的焦刚毛(sn)…”相关的问题
 得到了6个独立诱导而来的5突变的纯合子,每个都有AABB和aabb两种组成,将这6个纯合子互交以研究s基因的互补。结果如下表(表示野生型眼,s表示star眼,二者都是F1的表型):
得到了6个独立诱导而来的5突变的纯合子,每个都有AABB和aabb两种组成,将这6个纯合子互交以研究s基因的互补。结果如下表(表示野生型眼,s表示star眼,二者都是F1的表型):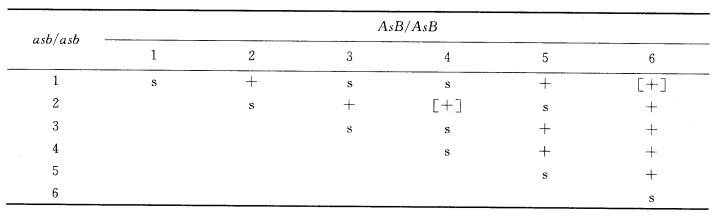 (1)在s位置有多少个顺反子?每个顺反子内有哪些突变位点? (2)使括号中的杂合子产生配子,在这两种情况中都发现了s+重组配子。检验了配子中两侧标记的组成,均为aB。给出这些顺反子相对Aa和Bb座位的顺序。
(1)在s位置有多少个顺反子?每个顺反子内有哪些突变位点? (2)使括号中的杂合子产生配子,在这两种情况中都发现了s+重组配子。检验了配子中两侧标记的组成,均为aB。给出这些顺反子相对Aa和Bb座位的顺序。
.jpg) 计算平均值、方差以及标准差。
计算平均值、方差以及标准差。.jpg)
.jpg) 解释上述结果。
解释上述结果。















