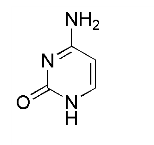
题目内容
(请给出正确答案)
题目内容
(请给出正确答案)
[主观题]
如果将纯化的大肠杆菌RNA聚合酶全酶(全酶中包含有δ亚基)在四种必需的核苷三磷酸存在的条件下加入到DNA,就合
如果将纯化的大肠杆菌RNA聚合酶全酶(全酶中包含有δ亚基)在四种必需的核苷三磷酸存在的条件下加入到DNA,就合成RNA。对所得到的RNA进行长度分析,表明mRNA分子的长度分布范围很广,且长度的分布是连续的。如果加入少量的细胞抽提液(已证实不含有核酸酶),则观察到长度不连续的较小的片段,试问抽提液中存在什么?
查看答案

 如果结果不匹配,请 联系老师 获取答案
如果结果不匹配,请 联系老师 获取答案

 更多“如果将纯化的大肠杆菌RNA聚合酶全酶(全酶中包含有δ亚基)在…”相关的问题
更多“如果将纯化的大肠杆菌RNA聚合酶全酶(全酶中包含有δ亚基)在…”相关的问题